
 Roger Abrantes
Roger Abrantes

The Illusive Free Will

The Illusive Free Will
—Do We Think Before We Act, or Do We Act Before We Think?
“Freedom is the individual’s capacity to know that he is the determined one, to pause between stimulus and response and thus to throw his weight, however slight it may be, on the side of one particular response among several possible ones.” This quote is from page 103 (of which I give you hereby a photo) of Lloyd-Jones, E., Westervelt, E. M. (1963) Behavioral science and guidance: proposals and perspectives. New York: Bureau of Publications, Teachers College, Columbia University.

We find this text frequently misattributed, misquoted, modified, and adulterated to suit various purposes and agendas on today’s overwhelming social media, where everybody knows everything—and nobody knows nothing (as they say).
Let’s take a breather for a moment, ponder, and analyze what’s behind the (original) quote without the social media’s razzle-dazzle.
There is a space—a tangible time gap—between the moment of an event and our reaction that we might use productively if we intentionally allow it to be there and engage it critically. That is, in other words, what the author means. It is an enticing proposition based on solid research from the previous decades.
The question of free will is a millennia-long one—in which neurophysiology only recently entered the fray. Theories of free-will focus on two fundamental questions: its possibility and nature. Some define free will as the capacity to make choices undetermined by past events. Determinism, on the other hand, sustains that only one sequence of events is possible and is inconsistent with free will. In contrast, compatibilism maintains that free will is consistent with determinism.
For over twenty years, experiments have suggested that, unbeknownst to us, a substantial part of mental processing occurs unconsciously, i.e., even before we know we plan to act. When we become aware of the brain’s actions, we ponder and mistakenly believe our intentions have caused them. “Your decisions are strongly prepared by brain activity. By the time consciousness kicks in, most of the work has already been done,” according to Haynes (2013).
Many processes in the brain occur automatically and without involving our consciousness. That is a defensive mechanism preventing our minds from being overloaded by basic routine chores.
We assume our conscious mind makes decisions. Our current findings question this. Researchers—using fMRI brain scans—could predict participants’ decisions up to seven seconds before the subjects had consciously made them.
If we decide before we are even aware of it, then the question is what mechanism decides for us. The prevailing view in neuroscience is that consciousness is an emergent neural phenomenon. The firing of the brain’s neurons results in consciousness and the sensation of free will or intentional action. These findings (Libet 1985) may not surprise neuroscientists who believe consciousness arises from brain activity (rather than brain activity originating from consciousness) since they view the conscious experience of free will as an emergent phenomenon of brain activity.
Researchers conclude that cerebral initiation of a spontaneous voluntary act begins unconsciously. However, within about 150 ms after the precise, conscious purpose emerges, we can still deliberately control the ultimate decision to act. Subjects can “veto” motor function for roughly 100–200 ms before a set time to act. That is the gap, the pause, so quoted and misquoted.
Whether that gap suffices to overcome the centuries-long free will quandary is highly arguable.
________________
References
- Dennett, DC (2003). Freedom Evolves. Viking Press.
- Fischer, JM & Ravizza, M (1998). Responsibility and Control: A Theory of Moral Responsibility. Cambridge University Press.
- Haynes, J-D (2013). World.Minds: Do We Have Free Will? (Charité Berlin).
- Libet, B (1985). Unconscious cerebral initiative and the role of conscious will involuntary action. Behavioral and Brain Sciences, 8(4), 529-539. doi:10.1017/S0140525X00044903.
- Park, H-D et al. (2020). Breathing is coupled with voluntary action and the cortical readiness potential. Nature Communications, 2020; 11 (1) DOI: 10.1038/s41467-019-13967-9.
- Pereboom, D (2001). Living Without Free Will. Cambridge University Press.
- Soon, C, Brass, M, Heinze, HJ, et al. (2008). Unconscious determinants of free decisions in the human brain. Nat Neurosci 11, 543–545. https://doi.org/10.1038/nn.2112
- Soon, CS, He, AX, Bode, S, Haynes, J-D (2013). Decoding abstract intentions, Proceedings of the National Academy of Sciences. March.
- Strawson, GJ (1994). The impossibility of moral responsibility. Philosophical Studies 75 (1-2):5-24.
Photo: Painting “The Illusion of Free Will (2016) by Daniel Stroup.
Featured Course of the Week
Ethology Ethology studies animal behavior in its natural environment. It is one of the fundamental courses in your curriculum. A reliable knowledge of animal behavior is the basis to create a satisfying relationship with any animal we train.
Featured Price: € 168.00 € 98.00

Ethology is the study of animal behavior in its natural environment. This online course, created by Ethologist Roger Abrantes, is unquestionably one of your curriculum’s most fundamental courses. Without reliable knowledge about animal behavior, we can’t expect to develop gratifying relationships with our pets or the animals under our care. Some of this course’s content is at an advanced level and requires increased dedication on your part. However, your expanded insight into the wondrous world of animal behavior will be a plentiful reward for your efforts.
The ethology online course comprises ten lessons, each with a quiz, and an ultimate test that awards you a diploma once completed. Included in the course is the beautiful flip-page book on which the course is based.
A Dog Is Not a Wolf—Is It?

The question has popped up several times in discussions on social media, with one faction adamant that wolves and dogs have nothing or very little to do with one another. As a result, we should not make comparisons between them.
As to how similar and different wolves and dogs are, I gave a lecture, “Wolf and Dog— A Comparative Study,” for the first time at the UNAM, Cuautitlán, Mexico, on May 2, 2017, covering the topic from (of course) a strictly scientific point of view. Having been studying wolves, dogs, and related canids for almost all of my professional career, I’ve collected plenty of evidence to pass judgment on the question.
It’s essential for us, as true behavioral sciences students, to stick to facts and sound argumentation. Therefore, I will give you here some points to help you evaluate the debate about wolves and dogs that we’ve seen on the internet, one dominated by hidden political agendas, emotional outbursts, and old wives’ tales. These facts will help you conduct a debate based on the currently available evidence and form an informed opinion on the topic.
Let me remind you that the term ‘compare’ in science refers to the discovery of similarities and differences. Both are equally fascinating and may supply us with valuable knowledge.
The following is only a short sample of the many more comparative facts we have. Please see the reference list.
● The dog family, Canidae, diverged from other carnivore families 50 to 60 million years ago. The family, comprising 34 extant species, shows a wide range of chromosome morphologies. The diploid chromosome number varies from 2n=36 (mainly metacentric autosomes) in the red fox, Vulpes vulpes, to 2n:78 (with all autosomes being acrocentric) in the domestic dog and various wolf-like canids such as the gray wolf, Canis lupus lupus. The chromosomal rearrangements in the different species help us deduce the group’s phylogenetic history (Wayne et al. 1987).
● Mitochondrial DNA sequences also reveal the Canidae’s evolution and the origins of the domestic dog (Wayne 1993). The results show that the domestic dog, Canis lupus familiaris is an extremely close relative to the gray wolf, with as little as 0.2% variation in mitochondrial DNA sequence between the two. That contrasts with the 4% variation in mitochondrial sequences between gray wolves and their nearest wild relative, the coyote, Canis latrans.
● Whole genome sequencing indicates that the dog (Canis lupus familiaris), the gray wolf (Canis lupus lupus), and the extinct Taymyr wolf diverged at around the same time 27,000–40,000 years ago (Callaway, 2015).
● Wolves and dogs differ by 0.2 to one percent, using the wolf-coyote time scale. That suggests they parted company about 135,000 years ago (based on samples from 140 dogs of 67 breeds and 162 wolves; see Robert Wayne, UCLA in Budiansky, S. 1999. The Atlantic Monthly; July 1999; The Truth About Dogs; Volume 284, No. 1; page 39-53).
● Canis lupus familiaris: 88% of its behavior is equal to the behavior of the Canis lupus lupus or slightly modified (Zimen, 1992; Feddersen-Petersen, 2004).
● Domestication seems to have caused a reduction in cooperative tendencies in dogs, mainly because of the significantly reduced cooperative breeding and hunting in dogs compared to wolves (Boitani & Ciucci, 1995; Coppinger & Coppinger, 2001; Kubinyi et al., 2007; Miklósi, 2007a; Range et al., 2009; Brauer et al., 2013).
● Unlike wolves, free-ranging dogs have a primarily promiscuous mating system (Daniels, 1983; Ghosh et al., 1984; Boitani et al., 1995; Pal et al., 1999; Pal, 2003). They rarely form monogamous pairs (for exceptions, see Gipson, 1983; Pal, 2005). Group members, other than their mother, rarely feed puppies (Macdonald & Carr, 1995; Boitani et al., 1995; Lord et al., 2013).
● Based on recordings of the social behavior of approximately 200 canines, including ritualized and non-ritualized forms of agonistic behaviors, play, and other affiliative behaviors, Feddersen-Petersen (2007) suggested that—contrasting to wolves—dogs have difficulties in cooperating even in a simple manner of just doing things together.
● Dog puppies show higher levels of aggressive behavior than wolf cubs (Feddersen-Petersen, 1991).
● In general, the ranking order (hierarchy) is a lot steeper in dogs than in wolves, resulting in a considerable social distance between the high-ranking animal(s) and the rest of the group (Feddersen-Petersen 1991).
In dogs, agonistic interactions often reach high aggression levels because dogs lack most strategies that wolves commonly use to solve conflicts, such as pacifying and inhibiting their opponents (Feddersen-Petersen 1991).
● Studies of free-ranging dogs assessed hierarchies using the same behavioral patterns described for wolves, i.e., submissive gestures, dominance displays, and aggressive behavior (Cafazzo et al., 2010).
● As in wolves, submissive gestures had a higher directional frequency within dyads (van Hoof & Wensing, 1987; Cafazzo et al., 2010).
● Results of studies suggest that the structure of the dog packs was relatively similar to that of wolf family packs, in which the ranking order is based on age, and males tend to dominate females within a given age class (van Hoof & Wensing, 1987; Mech, 1999; Packard, 2003).
● In small wolf family packs, both the breeding male and the breeding female share leadership (Mech, 2000).
● However, in large wolf family packs containing multiple sexually mature individuals, the higher-ranking breeders usually lead movements during 60%–90% of travel time (Peterson et al., 2002), meaning that in a non-negligible minority of cases, subordinate offspring can also provide leadership.
● Leadership was shared among group members in the studied dog packs, although not equally (Bonanni et al., 2010). Although every dog of at least one year of age could sometimes behave as a leader, each pack contained a limited number of ‘habitual leaders’ (i.e., individuals who behaved more frequently as leaders than they behaved as followers).
● Old and high-ranking individuals mainly provide leadership in the studied population’s free-ranging dogs (note that age and rank are positively correlated). This pattern seems to be relatively similar to that found in wolf family packs, in which parents lead activities (Mech, 2000; Peterson et al., 2002).
● In dogs, but not wolves, high-ranking animals show more aggression than lower-ranking ones about monopolizing the food (Ritter et al., 2012).
● Wolves are more tolerant than dogs during feeding competitions. Dogs develop a ‘steeper’ hierarchy that inhibits aggression directed towards higher-ranking group members (Ritter et al., 2012).
● Conclusion: Wolf and dog are quite similar in anatomy and physiology as to behavior, in some aspects more than others. See also the video “A Dog Is Not a Wolf—Is It?”
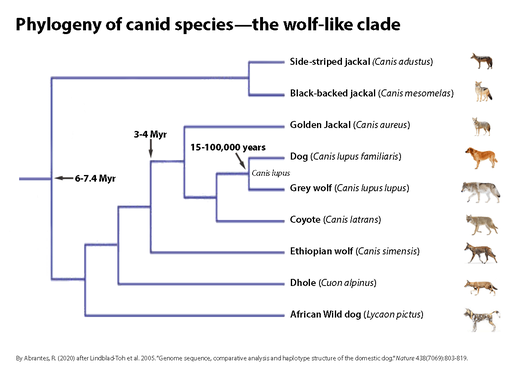
Phylogeny of canid species: the wolf-like clade. The phylogenetic tree is based on 15 kb of exon and intron sequence. The tree shown was constructed using maximum parsimony as the optimality criterion and is the single most parsimonious tree. Species names are represented with corresponding illustrations. Divergence time, in millions of years (Myr), is indicated for three nodes (Figure from Nicole Stange-Thomann, adapted by Roger Abrantes).
_____________
Note: For comparison, humans (Homo sapiens) and chimpanzees (Pan troglodytes) last shared a common ancestor ∼5-7 million years ago (Mya). The two species differ in DNA by ~4%. (Chen and Li 2001; Brunet et al. 2002; Varki, A., & Altheide, T. K. 2005). The complete mitochondrial DNA (mtDNA) molecules of Homo and the common chimpanzee were sequenced. The nucleotide difference between the entire human and chimpanzee sequences is 8.9%. The difference between the control regions of the two sequences is 13.9% and 8.5% between the remaining portions of the sequences (Arnason et al. 1996).
_____________
References
Abrantes, R. (1997). The Evolution of Canine Social Behavior. Wakan Tanka Publishers. ISBN: 0-9660484-1-5.
Arnason U, Xu X, Gullberg A. (1996). Comparison between the complete mitochondrial DNA sequences of Homo and the common chimpanzee based on nonchimeric sequences. J Mol Evol. 1996 Feb;42(2):145-52. doi: 10.1007/BF02198840. PMID: 8919866.
Boitani L. (1983). Wolf and dog competition in Italy. Acta Zoologica Fennica. 1983;174:259-264.
Boitani, L. & Ciucci, P. (1995) Comparative social ecology of feral dogs and wolves, Ethology Ecology & Evolution, 7:1, 49-72, DOI: 10.1080/08927014.1995.9522969.
Boitani L., Ciucci P., Ortolani A. (2007). Behaviour and social ecology of free-ranging dogs. In: Jensen P., ed. The behavioural biology of dogs. Wallingford, UK: CAB International; 2007:147-165.
Bonanni R. (2008). Cooperation, leadership and numerical assessment of opponents in conflicts between groups of feral dogs. Italy: Ph.D. thesis, University of Parma; 2008.
Brunet, M., Guy, F., Pilbeam, D., Mackaye, H.T., Likius, A., Ahounta, D., Beauvilain, A., Blondel, C., Bocherens, H., Boisserie, J.R., et al. (2002). A new hominid from the Upper Miocene of Chad, Central Africa. Nature 418: 145-151.
Bräuer, J., Bös, M., Call, J., & Tomasello, M. (2013). Domestic dogs (Canis familiaris) coordinate their actions in a problem-solving task. Animal cognition, 16(2), 273–285. https://doi.org/10.1007/s10071-012-0571
Callaway, E. (2015) Ancient wolf genome pushes back dawn of the dog. Nature. https://doi.org/10.1038/nature.2015.17607
Cafazzo S., Valsecchi P., Bonanni R., Natoli E. (2010). Dominance in relation to age, sex and competitive contexts in a group of free-ranging domestic dogs. Behav. Ecol. 2010;21:443-455.
Chen, F.C. and Li, W.H. (2001). Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees. Am. J. Hum. Genet. 68: 444-456.
Coppinger, R. & Coppinger, L. (2001). Dogs-A Startling New Understanding of Canine Origin, Behavior & Evolution. Bibliovault OAI Repository, the University of Chicago Press.
Daniels, T.J. (1983). The social organization of free-ranging urban dogs. II. estrous groups and the mating system, Applied Animal Ethology, Volume 10, Issue 4, 1983, ISSN 0304-3762, https://doi.org/10.1016/0304-3762(83)90185-2.
Feddersen-Petersen D. (1991). Verhaltensstörungen bei Hunden—Versuch ihrer Klassifizierung [Behavior disorders in dogs–study of their classification]. DTW. Deutsche tierarztliche Wochenschrift, 98(1), 15–19.
Feddersen-Petersen, D. (2004). Hundepsychologie. Sozialverhalten und Wesen. Emotionen und Individualität. 4. Auflage. Franckh-Kosmos. ISBN: 978-3-440-13785-7.
Feddersen-Petersen D.U. (2007) Social behaviour of dogs and related canids. In: Jensen P., ed. The behavioural biology of dogs. Wallingford, UK: CAB International; 2007:105-119.
Ghosh B., Choudhuri D.K., Pal B. (1984). Some aspects of sexual behaviour of stray dogs, Canis familiaris. Appl. Anim. Behav. Sci. 1984:13:113-127.
Gipson P.S. (1983) Evaluations of behavior of feral dogs in interior Alaska, with control implications. Vertebrate Pest Control Manag. Mater. 4th Symp. Am. Soc. Testing Mater. 1983;4:285-294.
Kubinyi, E., Virányi, Z., Miklosi, A. (2007). Comparative Social Cognition: From wolf and dog to humans. Comparative Cognition & Behavior Reviews. 2. 10.3819/ccbr.2008.20002.
Lindblad-Toh, K., Wade, C.M., Mikkelsen, T.S., Karlsson, E.K. (2005) Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 438(7069):803-819. DOI: 10.1038/nature04338.
Lord K., Feinstein M., Smith B., Coppinger R. (2013). Variation in reproductive traits of members of the genus Canis with special attention to the domestic dog (Canis familiaris). Behav. Processes. 2013;92:131-142.
Macdonald D.W., Carr G.M. (1995). Variation in dog society: between resource dispersion and social flux. In: Serpell J., ed. The domestic dog: its evolution, behaviour and interactions with people. Cambridge: Cambridge University Press; 1995:199-216.
Mech D. (1970). The Wolf: The Ecology and Behaviour of an Endangered Species. Garden City, NY: Natural History Press.
Mech L.D. (1999). Alpha status, dominance, and division of labor in wolf packs. Can. J. Zool. 1999;77:1196-1203.
Mech L.D. (2000). Leadership in wolf, Canis lupus, packs. Can. Field-Natural .2000;114:259-263.
Mech L. D., Boitani L. (2003). Wolf social ecology, in Wolves: Behavior, Ecology, and Conservation, eds Mech L. D., Boitani L. (Chicago; London: The University of Chicago Press; ), 1–35.
Miklosi, A. (2008). Dog Behaviour, Evolution, and Cognition. Dog Behaviour, Evolution, and Cognition. 1-304. 10.1093/acprof:oso/9780199295852.001.0001.
Packard J.M. (2003). Wolf behavior: reproductive, social and intelligent. In: Mech L.D., Boitani L., eds. Wolves: behavior, ecology, and conservation. Chicago, IL, and London: University of Chicago Press; 2003:35-65.
Pal, S.K., Ghosh B., Roy S. (1998). Agonistic behaviour of free-ranging dogs (Canis familiaris) in relation to season, sex and age. Appl. Anim. Behav. Sci. 1998;59:331-348.
Pal, S.K., Ghosh B., Roy S. (1999). Inter- and intrasexual behaviour of free-ranging dogs (Canis familiaris). Appl. Anim. Behav. Sci. 1999;62:267-278.
Pal, S.K. (2001). Population ecology of free-ranging urban dogs in West Bengal, India. Acta Theriol. 2001;46:69-78.
Pal, S.K. (2003). Reproductive behaviour of free-ranging rural dogs in West Bengal, India. Acta Theriol. 2003;48:271-281.
Pal, S.K. (2005). Parental care in free-ranging dogs, Canis familiaris. Appl. Anim. Behav. Sci. 2005;90:31-47.
Peterson, O., Jacobs A.K., Drummer I.D., Mech L.D., Smith D.W. (2002). Leadership behavior in relation to dominance and reproductive status in gray wolves, Canis lupus. Can. J. Zool. 2002:80:1405-1412.
Range F., Horn L., Bugnyar T., Gajdon G. K., Huber L. (2009). Social attention in keas, dogs, and human children. Anim. Cogn. 12, 181–192. 10.1007/s10071-008-0181-0.
Range, F., & Virányi, Z. (2015). Tracking the evolutionary origins of dog-human cooperation: the “Canine Cooperation Hypothesis.” Frontiers in psychology, 5, 1582. https://doi.org/10.3389/fpsyg.2014.01582.
Ritter, C., Viranyi, Z., Range, F., (2012). Who is more tolerant? Cofeeding in pairs of pack-living dogs (Canis familiaris) and wolves (Canis lupus). Third International Canine Science Forum.
van Hoof J.A.R.A.M., Wensing J.A.B. (1987). Dominance and its behavioural measure in a captive wolf pack. In: Frank H.W., ed. Man and wolf. Dordrecht, Olanda (Netherlands): Junk Publishers; 1987:219-252.
Varki, A., & Altheide, T. K. (2005). Comparing the human and chimpanzee genomes: Searching for needles in a haystack. Comparing the Human and Chimpanzee Genomes: Searching for Needles in a Haystack; genome.cshlp.org. https://genome.cshlp.org/content/15/12/1746.full#ref-10.
Vilà, C., Savolainen, P., Maldonado, J. E., Amorim, I. R., Rice, J. E., Honeycutt, R. L., Crandall, K. A., Lundeberg, J., & Wayne, R. K. (1997). Multiple and Ancient Origins of the Domestic Dog. Science, 276(5319), 1687–1689. http://www.jstor.org/stable/2892656
Wayne R. K. (1993). Molecular evolution of the dog family. Trends in genetics : TIG, 9(6), 218–224. https://doi.org/10.1016/0168-9525(93)90122-x
Wayne, R. K., Geffen, E., Girman, D. J., Koepfli, K. P., Lau, L. M., & Marshall, C. R. (1997). Molecular Systematics of the Canidae. Systematic Biology, 46(4), 622–653. https://doi.org/10.2307/2413498.
Wayne, R. K., Nash, W. G., & O’Brien, S. J. (2008). Chromosomal evolution of the Canidae. Cytogenetics and Cell Genetics 1987, Vol. 44, No. 2-3 – Karger Publishers.
Zimen, E. (1992). Der Hund – Abstammung, Verhalten, Mensch und Hund. Goldmann, 1992. ISBN-10: 3442123976.
Featured Course of the Week
Ethology Ethology studies animal behavior in its natural environment. It is one of the fundamental courses in your curriculum. A reliable knowledge of animal behavior is the basis to create a satisfying relationship with any animal we train.
Featured Price: € 168.00 € 98.00
A Girl and a Wild Mustang—or “There Are Many ‘Right’ Ways to Train Animals”

A Girl and a Wild Mustang
—or “There Are Many Right Ways to Train Animals”
“A Girl and a Wild Mustang” by Elsa Sinclair is a beautiful documentary—and yes, I’m biased.
I’ll explain why I find it beautiful and why I’m biased in a while. See, these days a tendency prevails in claiming there’s only one right way for us to interact with our animals. If you adhere to that idea, I fear I have bad news for you. There isn’t only one way to train animals; there are many. Let’s start from the beginning.
If you know a bit about evolutionary biology, you’ll know there are countless more ways to be dead than alive. Once we are alive, there are many ways of living, depending on anatomical, physiological, and environmental constraints. Once we’re over that, we are what we are, based on genetics, epigenetics, early imprinting, education, adaptation, goals, and circumstances.
It’s no surprise we differ from one another, no two humans, dogs, horses, or jellyfish being clones of each other.
In humans, the values, morals, and ethics we acquire throughout life accentuate peculiarities. Perhaps they do also in some other species (the evolutionary biologist suspects); only we don’t know.
Like/don’t like (the Facebook syndrome, as I coined the phenomenon) expresses our normative side (emotions and values) more than their descriptive counterparts (reason and facts). They reveal the radical and inescapable subjectivism and singularity of any conscious living organism—a condition Oriental philosophies heroically attempt to elude. We judge as we find suitable and as we can. We reason, but I’ll argue that normative judgments have the better of their descriptive analogs—at least for the human species at large. Perhaps once that gave us an evolutionarily advantageous edge.
Because we act according to our norms and not de facto facts, disagreement is expectable. Compound that with a good dose of faulty logic—the standard in human reasoning—and the probability of dissent moves one notch up from expectable to inevitable.
White is the safest color for a car, yet silver is the most popular. Why? Because car buyers chose by emotion, not reason. There’s nothing wrong with that (from a logical perspective) as long as we don’t claim that (A) safety is a concern of ours, and (B) color does not matter. Why is that so? Because, although the argument is valid, it is unsound.
The validity of an argument does not depend on its premisses being true, but its soundness does. A deductive argument is sound if and only if it is both valid, and all of its premises are true. In our example above, (B) is false, and the argument is, therefore, unsound.
There are many ways to train animals, depending on our knowledge, skills, goals, and preferences. Say, we want to establish a reciprocal, beneficial relationship with our dog or horse. We may choose any method we want: reinforcers only, combining reinforcers and inhibitors, a particular lead/leash, food or no food. Different approaches can be equally good. What we cannot do—provided we want to stay rational—is to justify our actions based on values with false facts.
What science tells us, does not oblige us to adopt any one method. Science doesn’t say a particular method is unconditionally better than another. It says that under specific circumstances A rates over B and B over C with a certain probability. A may be better for speed, B for reliability, and C for individual comfort. I choose, you choose, we choose what suits each of us best. We may want to go for speed, reliability, or what feels most comfortable for us. As long as we do not deny the facts science gives us, there’s nothing wrong with any choice of ours. What is wrong (unsound argumentation, as we saw above) is to state that we choose a particular method because of its efficiency for a specific purpose despite science telling us otherwise.
For example, during my career, I have assisted many owners and their animals. Invariably, my first questions would be, “what is your goal?” and “what price are you willing to pay?” And by price, I didn’t mean bucks, dimes, and pennies, but deviating from your values if need be.
Gimmicks—leads, collars, halters, food treats—speed up training a dog or a horse. Using neither is slower, and it requires much more knowledge, skill, and empathy. Again, it depends on what you want.
Students and clients often ask me which lead I recommend or prefer. My answer is always the same. I endorse none, but I can tell you what studies (not opinions) say about the effects of the various types—and you pick one, given your goals and principles.
If you ask me, I prefer none: no leads/leashes, collars, or any gimmicks. For me, the primary in any relationship with an animal (human or not) is the connection per se. I want it to be one between two free agents: no constraints besides the inevitable of having neighbors and having to share the world with others. That’s me, and I don’t expect you to think likewise—and I wouldn’t dream of imposing my view on you.
Given, I could achieve faster and more accurate and reliable results using other methods. Denying that is out of the question for it would compromise my intellectual integrity, which I am utterly incapable of doing. Blame my genetic constitution, early upbringing, education, and fortuitous circumstances.
Therefore, I’m willing to pay the price of sticking to my philosophy of life (and being slower, less accurate, etc.)—perhaps because I find the payback worth it. As I write elsewhere, “[…] (I) enjoy the closeness, not the training, not (the) achievement, […] relating to the animal (I) face as a living organism, an equal, a creature (I) meet for a brief moment in space and time.” (Animal Training My Way, 2015).
And so, you make your choices, to which you are fully entitled, and I mine. We are equally right as long as we do not deny facts or use faulty logic in our reasoning—which takes us to “A Girl and a Wild Mustang.”
As to my preferred interaction with animals (humans too), I’d suggest you watch Elsa Sinclair’s “A Girl and a Wild Mustang.” Sinclair made her choices and proved her point. Incidentally, she also proved my case, better than I could have done it—for which I’m grateful. You must see this documentary.
Sinclair does not impose her thoughts on you, rather states her case. It’s a take it, or leave it. As I said at the start, it’s a beautiful movie, and I am biased. My most avid readers will know right away, barely 30 seconds into the movie, why I recommend it so dearly.
Let me give you a teaser by quoting Sinclair: “Have you ever wondered if a horse would let you ride her if there was no threat of punishment or encouragement of a reward such as food? Would a horse let you ride her simply because you had built a strong relationship?”
I wrote in my book Evolution (2014), “We humans can contemplate nothing without changing it. Yet, the most important, it seems to me, must be to understand, accept, and respect other living beings independently of species and race.”
That’s a normative statement, all right, and one you may beg to differ. I read, recently, a claim that respect being a human construct, does not exist for dogs. I’ll dispute vigorously that respect is a human construct. Sinclair’s movie is all about mutual respect between human and horse.
“We both have the right to live. I have the right to chase you, you have the right to fly,” the lion says to the gazelle. I suspect respect, as in acting as others have the same freedoms as we do, even though they may conflict with ours, is a fundamental law of nature. If so, this statement of mine, paradoxically enough, will cease to be normative and will become a premise for this universe’s sustainability.
That is something to ponder on.
Photo by Elsa Sinclair. Artwork by Anton Antonsen.
Featured Course of the Week
Ethology Ethology studies animal behavior in its natural environment. It is one of the fundamental courses in your curriculum. A reliable knowledge of animal behavior is the basis to create a satisfying relationship with any animal we train.
Featured Price: € 168.00 € 98.00
Learn more in our course Ethology and Behaviorism. Based on Roger Abrantes’ book “Animal Training My Way—The Merging of Ethology and Behaviorism,” this online course explains and teaches you how to create a stable and balanced relationship with any animal. It analyses the way we interact with our animals, combines the best of ethology and behaviorism and comes up with an innovative, yet simple and efficient approach to animal training. A state-of-the-art online course in four lessons including videos, a beautiful flip-pages book, and quizzes.

Psychology Rather Than Power—for the Umpteenth Time, Reinforcers Are Not Rewards

For the umpteenth time, a reinforcer is not a reward. When I hear “Force-Free” trainers say, “dogs like to work to earn rewards,”1 I suspect and fear they miss by a mile and a half the essence and function of reinforcers in learning theory (and so also of inhibitors).
I’m not splitting hairs. There is a crucial difference between “reinforcing a particular behavior” and “rewarding an individual.” I suspect ignorance hereof being as well the cause of the many incorrect statements on inhibitors2 from the “Radical Force-Free” camp (emphasis on radical).3
“Dogs like to work to earn rewards” reminds me somehow of the old days when I opposed the old school of military dog training. Not everything was hassle and squabble; we had our moments of pleasantries. After ten minutes of marching forth and back, the instructor would say, “And halt! Now, praise your dogs.” Yes, I went to that kind of dog training with my first dog. That was what we had by then.
I walked out in disgust and decided, there and then, my dog and I would train by ourselves and we would show them. We did.
I substituted praise with reinforcers, the real thing—not rewards,—including my ‘dygtig’4 and treats given at strategic points. I stopped using a leash and started using a lead. Leash jerks gave place to “No” immediately followed by “dygtig” when the dog, not me, corrected the mistake. That, my dog would undisputedly do because she visibly enjoyed being my “teammate.” I was a student of ethology and I knew about social canines, our domestic dog being one. Old Professor Lorenz’s words rang in my ears, “To understand an animal, first you have to become a partner,” and he knew better than anyone for he had done it with his goslings.
I signed up for the final “obedience” competition at the club, a hunting dog club run by real hunters, and we won with max points. That a young long-haired fellow in faded Levi’s and clogs had won created some agitation; and, as to add insult to injury, my dog was a little, only seven-months-old English Cocker Spaniel (a genuine one, not one of those oddballs we see in the US today), red and female, on top of that. Petrine was intelligent, beautiful, charming, a workaholic, and a sweety-pie—though I might be a tad biased.
Our performance generated some raised eyebrows and more humming than the establishment would have wished. At the prizes and punch social function, a few civilians asked me whispering whether I would help training their fidos (read companion dogs).
The following Saturday, we were training on a grass field across the road where I lived, now the local firemen’s station. That was 1982, the summer before my son Daniel was born; and that’s how dog training came into my life. I never planned it.
Two years later, in 1984, I wrote my first book, “Psychology Rather than Force,” with far too little experience but loads of good ideas including force-free, hands-free, reinforcement-based training with as few inhibitors as possible, and it even included a whistle (the precursor of the clicker).
I was positive dog training would change. It did, and the rest is history.
__________
Notes
1 – This is an actual quote from a document published on the internet by a confessed “Force-Free” trainer.
Note that Skinner writes about reinforcers and rewards, “The strengthening effect is missed, by the way, when reinforcers are called rewards. People are rewarded, but behavior is reinforced. If, as you walk along the street, you look down and find some money, and if money is reinforcing, you will tend to look down again for some time, but we should not say that you were rewarded for looking down. As the history of the word shows, reward implies compensation, something that offsets a sacrifice or loss, if only the expenditure of effort. We give heroes medals, students degrees, and famous people prizes, but those rewards are not directly contingent on what they have done, and it is generally felt that the rewards would not be deserved if they had been worked for.” (Skinner, 1986, p. 569).
2- In 2013, I suggested we changed punisher and derivatives to inhibitor and derivatives to avoid the moral and religious connotations of the former, particularly in the Latin languages, and to emphasize its function and use as a learning tool.
3 – “Radical Force-Free dog trainers” (also “radical positives”) is my denomination for those trainers adhering to the positive reinforcement-based or force-free movement, but having extreme views like claiming positive reinforcers are the only learning tool one needs, they never use aversive stimuli, one should never say “no,” everyone else but them is wrong, and other absurdities. Please do not confuse them with the non-radical positive or reinforcement-based dog trainers who are equally force-free but sensible, open-minded, prudent in their claims, and polite and considerate to otherwise thinkers.
4 – “Dygtig” is my preferred semi-conditioned sound reinforcer. It’s a Danish word meaning’ clever,’ ‘skilled.’ It has a good doggy sound.
References
Abrantes, R. 1984. Psykologi Fremfor Magt (Psychology Rather Than Force). Lupus Forlag.
Abrantes. R. 2013. The 20 Principles All Animal Trainers Must Know. Wakan Tanka Publishers.
Skinner, B. F. 1986. What is wrong with daily life in the Western world? American Psychologist, 41(5), 568-574. Retrieved Jun. 29, 2019.
Featured photo by Annemarie Abrantes.
Featured Course of the Week
Ethology Ethology studies animal behavior in its natural environment. It is one of the fundamental courses in your curriculum. A reliable knowledge of animal behavior is the basis to create a satisfying relationship with any animal we train.
Featured Price: € 168.00 € 98.00
Learn more in our course Ethology and Behaviorism. Based on Roger Abrantes’ book “Animal Training My Way—The Merging of Ethology and Behaviorism,” this online course explains and teaches you how to create a stable and balanced relationship with any animal. It analyses the way we interact with our animals, combines the best of ethology and behaviorism and comes up with an innovative, yet simple and efficient approach to animal training. A state-of-the-art online course in four lessons including videos, a beautiful flip-pages book, and quizzes.

English Writing Aids

I’ve tested several English writing aids, tools that improve our writing and grammar. We have come a long way since the first, primitive spell checkers.
“If you propose to speak, always ask yourself, is it true, is it necessary, is it kind?” To mirror our current reality, I took the liberty to update this Buddhist saying.
“If you propose to post, always ask yourself, is it true, is it necessary, is it kind—is it well written?”
The way you say what you say matters, language matters.
Grammarly and ProWritingAid are by far the best English writing aids. Accuracy is similar in both apps; none is perfect. Since they are prone to miss different issues, using both improves our documents.
Grammarly is faster to use. ProWritingAid gives you more help—and work as well—for it goes into more details. To write a 100% satisfactory document for ProWritingAid is no mean feat; but you must be prudent since several suggestions, even though they may lighten the writing, will change your style.
Both apps allow you to save your documents in their clouds, which is an excellent facility for storage and quick editing. Both have browser extensions that will help to improve your writing nearly anywhere on the web.
Writing aids are like GPS instructions. They get you to your destination when you don’t know the route, and the more you know, the better they will serve you.
Bottom-line: Both are excellent tools. Grammarly is better suited to blogs, posts, and short essays. ProWritingAid appeals more to the professional author aiming at near perfection. Combining them is the top choice. None turns the pro copy editor superfluous yet. I recommend both apps for native and non-native English speakers. Because one is a native speaker does not mean one doesn’t commit language blunders or cannot improve. Both apps will turn you into a better writer.
See also “Recipe for answering questions.”
To achieve such a document score, you must work hard, especially in longer documents. PWA will detect colloquialisms, cliches, too many glue-words, passive sentences, sentence structure and variation, and much more.

You can see your sentence length at a glance. Varying the length of sentences makes it for more engaging reading.
Use adverbs with moderation in creative writing and outside dialogues. I use one adverb in my text, which is OK. Hidden verbs make for more difficult reading. PWA warns you of such instances.
Passive sentences are unclear. Write who, what, and when in unequivocal terms.

Sticky sentences contain too many common words and are not interesting for the reader. The optimal percentage is up to 40%. PWA gives my text a thumbs down by 0.1% which means it is virtually OK. I could correct that but I choose not to do it for it would change the style or tone of my post.
An interesting note: PWA warns me (not shown here) that “virtually” may be unnecessary. PWA is right. I disregard it, though, to give you an example of the depth of the app’s analysis.

A dynamic vocabulary makes for compelling reading. Readability measures show you how you can expect your readers to understand your message. Flesch over 60 is a must when you address readers at large. When you write to educated readers, the reading ease score may be lower. If you expect your audience to be non-native English speakers, keep your writing close to Flesch 60. In this caption, the vocabulary is more dynamic than 85% of ProWritingAid users and the Flesh score is 61%.

Featured artwork by Anton Antonsen, photo by Roman Kosolapov.
Featured Course of the Week
Ethology Ethology studies animal behavior in its natural environment. It is one of the fundamental courses in your curriculum. A reliable knowledge of animal behavior is the basis to create a satisfying relationship with any animal we train.
Featured Price: € 168.00 € 98.00
Take a look at Ethology and Behaviorism. Based on Roger Abrantes’ book “Animal Training My Way—The Merging of Ethology and Behaviorism,” this online course explains and teaches you how to create a stable and balanced relationship with any animal. It analyses the way we interact with our animals, combines the best of ethology and behaviorism and comes up with an innovative, yet simple and efficient approach to animal training. A state-of-the-art online course in four lessons including videos, a beautiful flip-pages book, and quizzes.
Dogs Point North-South When They Pee and Poop

Dogs point North-South when they pee and poop. They use the Earth’s magnetic field when urinating and defecating, aligning their bodies in the N-S axis.
If I wrote this on April 1st, everyone would take that for an April Fool’s prank. It is not. It is the conclusion of a scientific project conducted by Hart et al. and involving researchers from the Faculty of Forestry and Wood Sciences, Czech University of Life Sciences, and the Faculty of Biology, University of Duisburg-Essen in Germany (Hart et al. 2013).
Dogs Register Small Variations in Earth’s Magnetic Field
The study concludes dogs (Canis lupus familiaris) register small variations in Earth’s magnetic field after examining the behavior of 70 individuals (28 males and 42 females) belonging to 37 breeds, collected by 37 dog owners/reporters.
The researchers collected data on alignment during defecation (n = 1,893 observations, 55 dogs) and urination (n = 5,582, 59 dogs) from December 2011 through July 2013 in the Czech Republic and in Germany.
Under calm magnetic field conditions, dogs preferred to defecate with their bodies aligned along the north-south axis, even when sometimes facing south. Dogs not only favored N-S but also avoid E-W.
The data shows that larger and faster changes in magnetic conditions result in a random distribution of body alignments, i.e., a lowering of the preferences and ceasing of the avoidances, which may result from the magnetic disturbance or the intentional shutdown of the magnetoreception mechanism.
To avert any bias, all dogs moved in a free-roaming environment, off-lead and not restricted by walls or roads that would influence their movements. The routes of walks changed to exclude or limit pseudo-replication caused by the dogs defecating and urinating in the same few places.
The researchers also excluded the sun, polarized light, and the wind as determining factors for the body alignment of the dogs.
No Differences Between Males and Females
The study found no differences in the alignment of males and females during defecation and of the latter during urination. They all assume similar postures during defecation and females’ urination. Urinating males showed slightly different preferences to the females’ choices. The male leg lifting posture, while urinating, could explain these discrepancies.
No Answer to Why Dogs Prefer the North-South Axis
We have no answer to why dogs prefer the north-south axis and avoid east-west. It may be intentional, in which case they must perceive the magnetic field with one of their senses (as a haptic stimulus), or maybe they feel more comfortable aligning them in a particular direction (controlled at the vegetative level).
New Perspectives
Earlier studies confirmed that the natural fluctuations of the Earth’s magnetic field may disturb orientation in birds, bees, whales, and even affect vegetative functions and behavior in humans.
Studies on the wolves’ (Canis lupus lupus) homing are inconclusive. We cannot rule out the possible influence of an inherent sense of direction. There may be critical periods during a wolf’s life during which specific elements of its environment may imprint on it. In a study, the simplest hypothesis that explained the movement of four wolves was responses to visual, olfactory, and auditory cues with the latter probably being the most important. The wolves seemed driven to return to familiar territory, using the strongest learned exogenous cues (Henshaw and Stephenson 1974). Magnetoreception could have been a homing aid.
The findings of Hart et al. open new perspectives on how organisms use magnetic fields for direction.
References
Begall, S., Malkemper, P., Burda, H. (2014) Magnetoreception in mammals. Advances in the Study of Behavior, 2014. pp 45-79.
Dimitrova, S., Stoilova, I., and Cholakov, I. (2004) Influence of local geomagnetic storms on arterial blood pressure. Bioelectromagnetics 2004, 25:408–414. https://doi.org/10.1002/bem.20009.
Hart V, Malkemper EP, Kušta T, Begall S, Nováková P, Hanzal V, Pleskač L, Ježek M, Policht R, Husinec V, Červený J, Burda H. (2013) Directional compass preference for landing in water birds. Frontiers Zool 2013, 10:38. https://doi.org/10.1186/1742-9994-10-38.
Hart, V. et al. (2013) Dogs are sensitive to small variations of the Earth’s magnetic field. Frontiers in Zoology 2013, 10:80.
Henshaw, R.E., and Stephenson, R.O. (1974) Homing in the gray wolf (Canis lupus). J Mammal 1974, 55:234–237.
Southern, W.E. (1978) Orientation Responses of Ring-Billed Gull Chicks: A Re-Evaluation. In Schmidt-Koenig K., Keeton W.T. (eds) Animal Migration, Navigation, and Homing. Proceedings in Life Sciences. Springer, Berlin, Heidelberg, pp 311-317.
Featured illustration by Anton Antonsen.
Featured Course of the Week
Ethology Ethology studies animal behavior in its natural environment. It is one of the fundamental courses in your curriculum. A reliable knowledge of animal behavior is the basis to create a satisfying relationship with any animal we train.
Featured Price: € 168.00 € 98.00
Learn more in our course Ethology. Ethology studies the behavior of animals in their natural environment. It is fundamental knowledge for the dedicated student of animal behavior as well as for any competent animal trainer. Roger Abrantes wrote the textbook included in the online course as a beautiful flip page book. Learn ethology from a leading ethologist.
Canine Epigenetics

Epigenetics [Greek epi– ἐπι- = above, over, on, in addition to] is the study of heritable gene functions that cause stable phenotypic variations without affecting the DNA sequence of the organism.
Conrad Waddington coined the term in 1942 before we knew that DNA was the molecular basis of genes. He proposed that genes are differentially turned on and off by another level of “epigenetic” processes to produce different cells in the developing embryo.
Epigenetics, Environment, Phenotypic Plasticity
Behavioral epigenetics studies the role of epigenetics in forming behavior. It seeks to explain how nurture may shape nature. It attempts to provide a framework for understanding how the environment may influence gene expression to produce individual differences in behavior.
However, we must be careful with the term ‘environment’. To a geneticist, the environment is everything that isn’t the cellular environment of the DNA. To a social scientist, the environment catches everything from parental care to the stock market climate. That the cellular environment might be essential for understanding gene expression does not imply that one’s housing conditions have a similar impact.
The environment influences particular changes in gene action. For example, in alligators and specific turtles, egg incubation temperature affects the gene expression defining the sex of the individual. In these cases, the state of the gene passes down through cell divisions within a single organism, but it resets in eggs or sperm most times, so it does not transfer between generations. Thus, the mechanism does not influence evolution. An epigenetic state must carry over into the progeny to have any evolutionary relevance. For instance, molecules that bind to DNA and transfer to the offspring partially control the coat color of mice, Rattus norvegicus.
We must not confuse ‘epigenetics’ with ‘phenotypic plasticity,’ i.e., the capacity of one genotype of producing different phenotypes depending on the environment.
Belyayev’s Experiments
Belyayev’s experimented with silver foxes, Vulpes vulpes, which he bred based on a selection for tameness. He tested the animals, gave them a tameness score, and placed them in one of three groups. By the 20th generation, 35 percent of the animals were in the higher class, the ‘elite’ group; and as of 2009, ‘elite foxes’ made up 70 to 80 percent of the population. In addition, the changes in the tame foxes over the generations were not only behavioral but also physiological.
Belayev didn’t prove the effect of epigenetics in domestication. He proved the probability of domestication having occurred through selective breeding. In his own words, “It seems possible that the high frequency of the star mutation is due to strong selection intentionally applied for behavior.” (Belyaev et al. 1981). His experiments are a unique resource for studying the genetics of domestication.
The question of epigenetics goes deeper. Genes controlling plasma glucocorticoids were probably the targets during selection for tameness and the effects showed at all levels from phenotypic parameters to the gene expression of the corticotropin-releasing hormone (CRH), proopiomelanocortin (POMC), and the glucocorticoid receptor (GCR). Trut et al. conclude that “It appears plausible that the phenotypic novelties in the experimental fox population could be due to changes in gene activity, largely in its epigenetic modification.” (Trut et al. 2009).
If environmental conditions prefer a gene expression which parents pass to their progeny, and which, in turn, is more environmental independent than in the previous generation, then we can talk of a genuine epigenetic effect. If it is not, then we are talking about phenotypic plasticity and not epigenetics.
Most epigenetic changes occur only within the course of one individual organism’s lifetime—and that’s it. They can, though, pass to the organism’s offspring (transgenerational epigenetic inheritance). Also, if gene inactivation occurs in a sperm or egg cell that results in fertilization, this epigenetic modification may transfer to the next generation.
Conclusions
Do epigenetics make Lamarck right and Darwin wrong? To answer that, we must determine whether usage or selection causes epigenetic effects. Evidence supports the latter. Thus, epigenetic factors are yet another source of heritable natural variation. Darwin would have appreciated it. In fact, in 1868 he cautiously proposed ‘Pangenesis’ to cover the possibility that acquired characters might transfer to the progeny (whereby gemmules passed from somatic to reproductive cells). Modern discoveries seem to confirm that Darwin was right again.
At the time of writing, I have not found conclusive studies showing the effect of epigenetics on canine (Canis lupus familiaris) behavior, although Belyayev’s and subsequent studies make it plausible. I would appreciate if my reader has relevant information that may clarify this topic.
In popular writings appealing to the broad public, epigenetics seems to be everything that is not in the genes. However, that is not the scientific view, one requirement being that an effect is only epigenetic if it impacts the evolution of a trait. Therefore, I would recommend prudence when analyzing any statement claiming ‘epigenetic’ effects. These days, the term ‘epigenetic’ (like ‘quantum’) is prone to arouse the fantasy of quacksalvers.
References
Belyayev, D.K., Ruvinsky, A.O., and Trut, L N. 1981. Inherited activation/inactivation of the star gene in foxes. Journal of Heredity, 72: 264-274.
Chandler, V.L. 2007. Paramutation: from maize to mice. Cell. 128 (4): 641–45. doi:10.1016/j.cell.2007.02.007. PMID 17320501.
Cimarelli, G., Virányi, Z., Turcsán B., Rónai, Z., Sasvári-Székely, M., Bánlaki, Z. 2017. Social Behavior of Pet Dogs Is Associated with Peripheral OXTR Methylation. Front Psychol. 2017 Apr 10;8:549. doi: 10.3389/fpsyg.2017.00549.
Darwin, C. R. 1868. The variation of animals and plants under domestication. London: John Murray. 1st edition, second issue. Volume 1.
Dias, B.G., & Ressler, K.J. 2014. Parental olfactory experience influences behavior and neural structure in subsequent generations. Nature Neuroscience, 17(1), 89-98. doi: 10.1038/nn.3594.
Dupont C., Armant D.R., Brenner, C.A. 2009. Epigenetics: definition, mechanisms and clinical perspective. Seminars in Reproductive Medicine. 27 (5): 351–57. doi:10.1055/s-0029-1237423. PMC 2791696. PMID 19711245.
Florean, C. 2014. Food that shapes you: how diet can change your epigenome. Science in School, Issue 28.
Hughes, V. 2014. Epigenetics: the sins of the father. Nature, 507, 22-24.
Kaplan, G. 2017. Why is my dog this way, does it matter if we know, and what can we do? IAABC journal.
Pörtt, D., Jung, C. 2017. Is dog domestication due to epigenetic modulation in brain? Dog Behavior. Vol 3, No 2 (2017). ISSN 2421-5678.
Trut, L.N. 1996. Sex ratio in silver foxes: effects of domestication and the star gene. Theor Appl Genet (1996)92:109-115. ISSN
1432-2242.
Trut, L., Oskina, I., and Kharlamova, A. 2009. Animal evolution during domestication: the domesticated fox as a model. Bioessays. 2009 Mar; 31(3): 349–360.
Featured image by Anton Antonsen, photo by Hitdelight.
Featured Course of the Week
Ethology Ethology studies animal behavior in its natural environment. It is one of the fundamental courses in your curriculum. A reliable knowledge of animal behavior is the basis to create a satisfying relationship with any animal we train.
Featured Price: € 168.00 € 98.00
Learn more in our course Ethology. Ethology studies the behavior of animals in their natural environment. It is fundamental knowledge for the dedicated student of animal behavior as well as for any competent animal trainer. Roger Abrantes wrote the textbook included in the online course as a beautiful flip page book. Learn ethology from a leading ethologist.
Do Animals Show Altruism?

Introduction |Etymology | Definition and Problems | The Altruism Paradox- Group selection—Wilson | Kin Selection—Maynard Smith | Reciprocal Altruism—Trivers | Genetic Selfishness—Dawkins | Phenotypic plasticity | An Example We All Know—Do Dogs Show Altruistic Behavior? | Conclusion and Perspectives
________
Altruism means unselfishness, selflessness, the regard for others as a principle (the opposite of egoism or selfishness). We all use the word in our daily language, feeling confident we know everything about altruism, its meaning, and implications, but do we? The concept poses many questions to biologists, philosophers, and other scientists. It has always done. For one, defining altruism isn’t as straightforward as the introductory paragraph above may seem to imply. Then, setting up a model where such behavior makes sense and can evolve without contradicting evolutionary theory has proven to be a real brain twister.
Etymology
Altruism derives from the French autrui = other people. Autrui developed from the Old French autre = other which itself comes from Latin alter = other. [1] Under the influence of alter, the French autrui produced the altrui- of the French altruisme and the English altruism. The English term has been in use since the mid-1800s. The translation of Auguste Comte (the French founder of the philosophical school of positivism), who coined the term for an antonym of egoism, [2] contributed to spreading its adoption in English.
Definitions and Problems
What is altruism? The vague definition of the term makes us doubt whether altruistic behavior is possible at all. Note that most synonyms of ‘altruism’ are not much more enlightening—also needing some explaining. If to be altruistic, the actor must get nothing at all in return for its actions, then we may question if any act qualifies as altruistic. Self-satisfaction always seems to follow a charitable act. Therefore, according to the theory of psychological egoism, we cannot describe sharing, helping, or sacrificing as altruistic. [3]
If we define altruism as unselfish conduct in the short term, i.e., without immediate benefit, then we can find more examples of such acts. One typical case is parental behavior. Parents do plenty for their progeny with no direct advantage to themselves. Female canine mothers, including our domestic dogs, Canis lupus familiaris, spend enough time and energy looking after their pups, feeding them, and teaching them various skills. [4] They regurgitate for them and defend them if they are in danger.
In the wild social canids, pack members also support caring for youngsters without immediate benefits. In jackals, Canis aureus—a group comprising father, mother, and cubs—when a juvenile is present (almost always, a sister to the newborn), she will help rear her younger sibling. [5].
However, note that some behavior we classify as altruistic may have a different and more plausible explanation. The case (in the Samburu National Park in Kenya) of a female lion adopting an oryx calf is more likely to result from a hormonal imbalance affecting its maternal instinct, than to altruism. [6]
Last, a problem of great concern, one that had preoccupied great minds in biology, is the ‘altruism paradox.’
Darwin well knew of that. He writes, “It is extremely doubtful whether the offspring of the more sympathetic and benevolent parents, or of those who were the most faithful to their comrades, would be reared in greater numbers than the children of selfish and treacherous parents belonging to the same tribe. He who was ready to sacrifice his life, as many a savage has been, rather than betray his comrades, would often leave no offspring to inherit his noble nature.” [7]
The Altruism Paradox
In explaining kin selection, evolutionary biologists imagine a gene that causes its bearer to behave altruistically toward other organisms and those without it to behave selfishly. “The altruists will be at a fitness disadvantage, so we should expect the altruistic gene to be eliminated from the population.” [8] That is, in a nutshell, the problem of altruism for the evolutionary scientist.
How can an altruistic gene, coding for the weaker altruistic phenotype, thrive in competition against a selfish one that codes for the fitter selfish phenotype? Hence, we have the altruism paradox. As we shall see, there might be a workaround this issue, but let us first review the traditional models.
Group Selection
Group selection is another proposed mechanism of evolution in which natural selection operates at the group level, instead of at the level of the individual [9][10].
Darwin, in “The Descent of Man” in 1871,[7] attempted to explain the evolution of human altruism as a selection process at the group level: “When two tribes of primeval man, living in the same country, came into competition, if (other things being equal) the one tribe included a great number of courageous, sympathetic and faithful members, who were always ready to warn each other of danger, to aid and defend each other, this tribe would succeed better and conquer the other.” [7]
Darwin’s explanation seems a tad non-Darwinian, thus revealing the magnitude of the problem.
A winner has no impact on evolution, per se, unless it has a better ‘Darwinian or inclusive fitness’ (the genetic contribution of an individual to the next generation’s gene pool relative to the average for the population). An altruist may win but if its ‘Darwinian and inclusive fitness’ is nil, altruism stops with it. If altruism survives, even though donors perish leaving no progeny, the survival of the fittest is not true for the fittest are those who leave the most copies of themselves in successive generations.
This issue has bothered many since Darwin, among them, Hamilton [11]. E. O. Wilson and D. S, Wilson write: ‘‘[…] something more than natural selection within single groups is required to explain how altruism and other group-advantageous traits evolve by natural selection.” [12]
For natural selection to favor altruism in a broader scenario, the ‘within-group’ disadvantage of the altruist must be offset by the ‘between-group’ advantage of the group including altruists. [13] ‘‘Cooperation is always vulnerable to exploitation by defectors; hence, the evolution of cooperation requires specific mechanisms, which allow natural selection to favor cooperation over defection.’’ [14][15]
For group selection to be viable, we must assume that the variation between groups is larger than the variation within groups. Since selection acts upon the phenotype, competition and selection can operate at all levels. Therefore, D. S. Wilson contends that “At all scales, there must be mechanisms that coordinate the right kinds of action and prevent disruptive forms of self-serving behavior at lower levels of social organization.”[16] He summarizes, “Selfishness beats altruism within groups. Altruistic groups beat selfish groups. Everything else is commentary.”[16]
As we shall see, not everyone agrees with that. ‘Between-group’ selection is possible, in principle, although it is weak compared to any which may happen ‘within-group’. Therefore, if we are to explain ‘for the good of the group’ behavior, then we must do it without group selection.
In fact, all models for explaining how cooperative and altruistic social behavior evolve, such as kin selection, reciprocity, and the selfish gene theory developed as alternatives to group selection.
Kin Selection
To explain altruism, we must find a way of natural selection to favor altruistic genes as in the theory of kin selection.[11] Kin selection is the evolutionary strategy that supports the reproductive success of an organism’s relatives at one’s cost. It is kin altruism based on inclusive fitness. Maynard Smith used the term ‘kin selection’ for the first time in 1964.[17]
Charles Darwin discussed this strategy in “The Origin of Species,” (1859)[18] arguing that a selection benefit to “the same stock” (kin) would allow the evolution of a trait while destroying an individual. Seventy years later, Fisher and Haldane figured out the mathematics of kin selection. [19][20] According to Maynard Smith, Haldane resumed the conclusion of his calculations by saying that, “he was prepared to lay down his life for eight cousins or two brothers.” [21]
Hamilton’s inclusive fitness rule states that kin selection increases the frequency of particular genes when the genetic relatedness of a recipient to a donor multiplied by the benefit to the recipient is greater than the reproductive cost to the donor.
rB>C
where r=the genetic relatedness of the recipient to the donor; B=the reproductive benefit gained by the recipient of the altruistic act; C=the reproductive cost to the donor.
Thus, kin selection is a special consequence of gene selection. The degree at which one should extend altruistic behavior toward others depends upon their coefficient of relationship. There are two ways to achieve this: (1) by kin recognition and/or (2) by living near one’s relatives (Hamilton 1964).
‘Kin selection’ is not the same as ‘group selection’ where a genetic trait may become widespread because it benefits the group as an entity.
Reciprocal Altruism
Reciprocal altruism is all behavior whereby a donor, with one act, reduces its fitness while increasing a recipient’s fitness, expecting subsequent payback. [22] Thus, the reduction is temporary. The mechanism is close to the “tit for tat” strategy in game theory.[23]
Reciprocal altruism, regarded as an instance of the prisoner’s dilemma, [24] is an evolutionary possibility if chances of meeting another reciprocal altruist are high enough, or if the game continues long enough.[22]
Reciprocal altruism, [22] is a possible way for natural selection to favor altruistic genes provided that:
(1) the individuals must have plenty of opportunities for reciprocation;
(2) they must be able to recognize each other as individuals;
(3) they must remember the obligations;
(4) and that they must be motivated to reciprocate.
Although this model seems to be evolutionarily unstable, evolutionary biologists found a way in which it would work.
There are striking parallels between altruistic behavior and exaggerated sexual ornaments. Both are costly in fitness and easy to detect, and both might be fitness signals turned evolutionarily stable by the handicap principle. The handicap principle suggests that honest communication is expensive to the signaler, therefore only affordable to special individuals. Receivers know that reliability must backup quality because lesser signallers couldn’t afford such extravagances. [25][26][27]
Then, we have the homogeneousness norm. Change in phenotype and functionality, caused by a non-silent mutation, will often stand out in a population. Thus, we can expect sexual individuals to prefer mates with the least number of unusual or minority features. As a result, and given enough time, a whole population will develop similar looks. In a similar way, the behavior repertoire of a population will become evolutionarily stable once it has developed as homogeneous as is the rule in most species. This includes any altruistic and cooperative features.[28]
Genetic Selfishness
When parents sacrifice themselves for their progeny, they are benefiting themselves since their offspring have 50% of their genes. Therefore, we can regard altruistic behavior as genetically selfish. That is Haldane’s extrapolation from ‘kin selection’.
For example, in wolves, Canis lupus lupus, [29] it pays off for each parent to sacrifice its life saving two of their progeny because this equals twice 50% of their own genes. However, such a calculation is more complicated than so if we take into account the cubs’ chances of survival without parental support. Giving their lives to save their youngsters when they are only one week old is a bad trade since they are unlikely to survive. In this instance, the best strategy is for the male and the female to protect themselves and keep the prospect of producing more offspring later.
This model explains why individuals sacrifice more for their progeny than for those of relatives or strangers, as we saw.[30] What the defenders of the selfish gene want to emphasize, besides agreeing with kin selection and inclusive fitness, is that the selection process happens at the gene level.[31][32]
In the genetic selfish model, the unit of replication is the gene, and the organism is the vehicle it uses and upon which selection acts directly. “Natural selection favours some genes rather than others not because of the nature of the genes themselves, but because of their consequences—their phenotypic effects.”[32]
Because genes are selfish, they will promote selfish behavior in the individuals they produce. ‘Selfish’ means, in this sense, to take care of itself as the first priority. Dawkins writes, “[…] gene selfishness will usually give rise to selfishness in individual behaviour. However, […] there are special circumstances in which a gene can achieve its own selfish goals best by fostering a limited form of altruism at the level of individual animals.”[32]
Gene selection soundly explains kin selection and eusociality. An organism acts altruistically, against its individual interests, because by supporting a related one to reproduce, genes help copies of themselves (or sequences with the same phenotypic effect) in other bodies to replicate. Thus, sometimes, ‘selfish’ actions by the genes lead to unselfish behavior by the organisms.
The survival of each gene, being replicators, depends on the survival of some others. Being a selfish gene does not imply that genes are entirely uncooperative. To be successful, a gene needs to cooperate with the other genes with which it shares a phenotype. Genes cooperate in building bodies because they all share the same exit route into the next generation. That’s their only way to survive. An organism, a body, is a vehicle for its genes, built up by a cooperative of genes.
Vehicles are important, but replicators are essential. Darwinian natural selection is still viable with no vehicles, only replicators, but not the other way around. In fact, when life began, there were probably no vehicles, only replicators.
A group is not a replicator because there is no ‘group pool’ (like there are ‘gene pools’). There is no metapopulation in which some groups are more successful than others at making replicas of themselves. And a group is not a vehicle because to qualify as one, all the genes in the same group would need to share the same exit route to the next group in the generational sequence.
Selection based upon groups is rare compared to selection on individuals, according to the selfish gene model. Researchers could not confirm simple interpretations of group selection, though more sophisticated ones proved to make accurate predictions in specific cases.[12]
E. O. Wilson writes that although the selfish-gene approach has been widely accepted “[…] Martin Nowak, Corina Tarnita, and I demonstrated that inclusive fitness theory, often called kin selection theory, is both mathematically and biologically incorrect.” He argues that group selection is a more realistic model of social evolution.[12][33[[34]
Dawkins rejects replacing ‘kin selection’ with ‘group selection.’ According to the selfish-gene model, viewing evolution as driven by the differential survival of whole groups of organisms is incoherent. He does not deny ‘group selection’. What he contends is that, even in the rare cases where it is not wrong, it is cumbersome, time-wasting, and distracting to what would otherwise be a straightforward understanding of what happens in natural selection.
Both Dawkins and Wilson agree that favorable genes are likely to prosper and replicate and that living in groups is helpful in some circumstances. The dispute arises mostly over definitions. They aim toward representing empirical facts with precision, but both use too broad definitions for ‘group,’ ‘group selection’ and ‘kin selection’. Thus, it becomes rather difficult to test their models.
We might find a consensus if we can simulate various evolutionary scenarios with narrower definitions, similar to what Markvoort et al. did for simulations of cellular group selection.[35]
A Workaround the Classic Approach
The altruism paradox, as we saw, goes down to the question: how can an altruistic gene, coding for the weaker altruistic phenotype, thrive in competition against a selfish one that codes for the fitter selfish phenotype?
Perhaps this is a confirmation that an answer cannot be better than the question it addresses. What if we are posing the wrong question and, hence, creating the paradox?
The thought of a distinct altruistic allele risking being overrun by a distinct selfish one is the basis for all the models we reviewed above.
As Dawkins writes, “‘cheat genes’ are spreading through the population while ‘sucker’ genes are driven to extinction.”[32] E. O. Wilson also summarized the problem in the same lines, “How might such a behavior evolve if the genes promoting it are at such a disadvantage in competition with genes that oppose it?” [36]
Now, suppose there are no two genotypes coding for the two competing phenotypes, selfishness and altruism. What if both phenotypes were due to one single genotype carrying both alternatives—no two distinct alleles? The Altruism Selfishness Plasticity (ASP) model suggests exactly that. Thus, let us for a moment set the prevailing Altruism Selfishness Allelomorphism (ASA) frame aside and explore what the ASP approach can add to the debate.
Altruistic and Selfish Phenotypes as Plastic Expressions of a Single Genotype
There is a way around the altruism paradox: to consider the altruistic and selfish phenotypes as plastic expressions of a single genotype. [37]
Phenotypic plasticity is the property of a genotype to produce different phenotypes in response to distinct environmental conditions.[38] There is nothing strange about that. We have known about phenotypic plasticity for quite a while. It denotes particular morphological and physiological changes in an organism—and behavior—as a response to a specific environment. Although biologists, to begin with, used the term to describe some developmental effects of a morphological character, they use it today to describe all phenotypic responses to environmental conditions, e.g., acclimatization and learning.[39]
In fact, we have plenty of data to support the ASP model. It is very plausible that no distinct genotype codes either the altruistic or the selfish phenotype, that they are rather plastic expressions of the same genotype, determined by particular environmental circumstances. And if this is true—the altruistic and selfish phenotypes being creations of a single genotype—then, they cannot be competing. To use Dawkins’ metaphor, the same genotype has the plasticity to express itself either as a ‘cheater’ or a ‘sucker’ depending on accidental contingencies.
Yakubu provides plenty of evidence in support of such a solution to the altruism paradox.[37] Let us review some of them.
The Social Hymenoptera
The social Hymenoptera are a favorite of biologists because they are highly social and haplodiploid. A honeybee, Apis mellifera, colony has three castes consisting of a queen, a few hundred males (drones), and thousands of non-reproductive females (workers). The queen’s role is to reproduce and the drone’s job is mating a queen. The workers, on the other hand, strive hard keeping the colony, foraging, and defending it from intruders. The reproductive queen is selfish, while the nonreproductive workers are perhaps the epitome of altruism.
Now, we can ask whether there are distinct alleles (genotypes) for altruism and selfishness in eusocial populations as the classical models of altruism presume. Evidence refutes that conclusively.
Whether a larva becomes a queen or a worker begins with where the egg is laid and continues with the received feeding.[40] The workers will nurse a larva in a queen cell with royal jelly.[41] On the other hand, if the larva is in a worker cell, they will feed it worker food. Amazingly enough, the former will become a queen and the latter a worker. We can say the eggs and the larva are totipotent [40] or multipotent (according to this author).
The honeybee provides us with even more staggering evidence. We can move eggs and larva from a queen cell to a worker cell, or the other way around, and if we do that within the first three days, they will develop as they are fed and as to where they reside.[40] We can thus infer that there is no genotype involved in the queen/worker distinction. Whether a bee becomes selfish (queen) or altruist (worker) depends only on environmental stimuli.
The caste differentiation happens via an epigenetic process where non-heritable factors contribute to gene expression. Queen and worker morphological forms both originate in the same genome. Royal jelly nourishment is the non-genetic determiner. The genes encoding the major royal jelly proteins present one of the clearest examples of a gene class acquiring new functions during the evolution of sociality.[42]
Researchers have sequenced the genome of the honeybee.[43] In fact, we know the particular genes in the honeybee whose differential expression results in queen and workers. Evans and Wheeler identified and characterized transcripts from seven genes that are expressed differently by the worker- and the queen-destined larvae at a critical point in their development. They are indeed genes with phenotypic plasticity.[44]
Altruistic Expression and Social Cues
Let us now consider non-eusocial social organisms. Trivers explained the Vampire bats altruistic behavior based on the ASA assumption.[22] However, we can explain that behavior as well without presuming the existence of distinct selfish/altruistic genotypes.
In vampire bats, hungry individuals often solicit food from those that are better fed. Sometimes, an individual obliges and regurgitates to a soliciting individual—and, sometimes, it steadfastly refuses to share food.
Trivers is right as to the reciprocity of this altruistic behavior. Whether a vampire bat shares blood depends on whether the solicitor has given the donor blood earlier or is likely to give blood to the donor later.[45] The same individual behaves selfish and altruistic depending on social circumstances. This conclusion is compatible with reciprocal altruism.[37] However, as genotypes do not change overnight, it is more plausible for the distinct phenotypes to be due to the plasticity of the same allele.
Male adult olive baboons, Papio anubis, help troupe members depending on whether he has earlier received help from those individuals or they are deemed likely of lending him help in the future.[46]
The behavioral strategy of efficient coercion also supports the plastic phenotypic deployment of a single genotype. In ten studied social insect species, social sanctions kept individuals altruistic in situations where they would have been selfish (Wenseleers and Ratnieks 2006).[47]
Altruists can become selfish. In white-fronted bee-eater, Meropsis bullockoides, young males do not set up nests because older males harass them. They become altruistic helpers instead. However, some of these helpers turn selfish once they build their own nests.[48]
In meerkat societies, the altruistic lower-ranking females become selfish if they attain a higher ranking, not only by breeding but also by killing the infants of other (by then) subordinate females.[49]
Social organisms may assume subordinate roles, not because of their altruistic genes, but because it is the best of the options, they have at the time.[50]
The reviewed examples seem to confirm the hypothesis of altruism being due to a single genotype with plastic phenotype options. An individual will behave altruistically under particular environmental conditions and will respond selfishly when those same cues are absent. As far as this author knows, no evidence has yet proved that only certain individuals can react altruistically or selfishly, given they act in the same scenario.[37]
An Example We All Know—Do Dogs Show Altruistic Behavior?
We know dogs for teaming together against rivals. That is widespread territorial behavior among pet dogs, strays and wild canids. They also defend their household, including humans and other animals. The females bring up and protect their pups at their own expense. These are all examples of behavior we can classify as altruistic. Even considering the difficulties inherent to the concept of altruism, we described, a prudent statement would be that dogs show reciprocal altruistic behavior under the right circumstances, as the ASP model describes.
Conclusion and Perspectives
The altruism paradox is perhaps among the most interesting in ethology (biology) and has, therefore, preoccupied many researchers. It poses highly pertinent questions to enhance our understanding of behavior, genetics and evolutionary theory. Scientists have proposed various models, some of which are more consensually accepted than others. Still, they have all contributed to increasing our global knowledge. The application of the ASP model to behavior promises perspectives far beyond the one we studied in this paper. Perhaps other behaviors that conventional approaches ascribe to distinct phenotypes are but phenotypic plastic expressions of a genotype.
References
[1] “Altruism.” Merriam-Webster. Merriam-Webster, Incorporated. 2017. Retrieved 2017-10-10.
[2] Altruism, Based on the Random House Dictionary (2017). Dictionary.com Unabridged. Random House, Inc. 2017. Retrieved 2017-10-10.
[3] Slote, M.A. 1964. An Empirical Basis for Psychological Egoism, Journal of Philosophy 61: 530-537. Retrieved: 2017-10-10.
[4] Fox, M.W. 1972. Behaviour of Wolves, Dogs, and Related Canids. Harper & Row. ISBN 978-0060113216.
[5] Rogers, L.J.; Kaplan, G.T. 2003. Spirit of the Wild Dog: The World of Wolves, Coyotes, Foxes, Jackals & Dingoes. Allen & Unwin. ISBN 1865086738.
[6] “An amazing True Story—1/24/2008“. The David Sheldrick Wildlife Trust. 2008. Retrieved 2017-10-10.
[7] Darwin, C. 1871. The Descent of Man, and Selection in Relation to Sex (1st ed. ed.). London: John Murray. ISBN 0-8014-2085-7.
[8] Okasha, S. 2009. Biological altruism. In: Zalta EN (ed) The Stanford encyclopedia of philosophy, winter 2009 edn.
[9] Wynne-Edwards, V.C. 1962. Animal Dispersion in Relation to Social Behaviour. Edinburgh: Oliver & Boyd.
[10] Lorenz, K. 1963. Das sogenannte Böse zur Naturgeschichte der Aggression. Verlag Dr. G Borotha-Schoeler (“So-Called Evil, Toward a Natural History of Aggression”).
[11] Hamilton, W.D. 1964. The genetical evolution of social behaviour, I and II. Journal of Theoretical Biology. 7: 1–16, 17–32. doi:10.1016/0022-5193(64)90038-4.
of sociobiology. Q Rev Biol 82:327–348.
[13] Taylor, C., Nowak, M.A. 2007. Transforming the dilemma. Evolution, 61(10):2281–2292.
[14] Allen, B., Traulsen, A., Tarnita, C.E., Nowak, M.A. 2012. How mutation affects evolutionary games on graphs. J Theor Biol 299:97–105.
[15] Nowak, M.A. 2012. Evolving cooperation. J Theor Biol 299:1–8.
[16] Wilson, D.S. 2015. Does Altruism Exist? Culture, Genes, and the Welfare of Others. Yale UP. ISBN: 0300189494.
[17] Maynard Smith, J. 1964. Group selection and kin selection. Nature. 201 (4924): 1145–1147. doi:10.1038/2011145a0.
[18] Darwin, C. 1859. The Origin of Species. Reprint: Modern Library, New York 1998.
[19] Fisher, R. 1930. The Genetical Theory of Natural Selection. The Clarendon Press Oxford. OCLC: 18500548.
[21] 1976 July 29, New Scientist, Section: Letters, Haldane, (Letter to the editor from J. Maynard Smith), Quote Page 247, Column 1 and 2, Published by New Science Publications, London; Now Published by Reed Business Information, UK.
[22] Trivers, R. L. 1971. The evolution of reciprocal altruism. Quarterly Review of Biology. 46: 35–57. doi:10.1086/406755.
[23] Rapoport, A. 1966, Two-Person Game Theory: The Essential Ideas. The University of Michigan Press. Ann Arbor, MI,
[24] Mérő, L. 1998. The Prisoner’s Dilemma. In: Moral Calculations. Springer, New York, NY. https://doi.org/10.1007/978-1-4612-1654-4_3.
[25] Fisher, R. 1930. The Genetical Theory of Natural Selection. The Clarendon Press Oxford. OCLC: 18500548.
[26] Zahavi, A. 1975. Mate selection—a selection for a handicap. Journal of Theoretical Biology. 53 (1): 205–214. PMID 1195756. doi:10.1016/0022-5193(75)90111-3.
[27] Zahavi, A. 1977. The cost of honesty (Further remarks on the handicap principle). Journal of Theoretical Biology. 67 (3): 603–605. doi:10.1016/0022-5193(77)90061-3.
[28] Koeslag, J.H. 1997. Sex, the Prisoner’s Dilemma Game, and the evolutionary inevitability of cooperation. Journal of Theoretical Biology. 189: 53–61. PMID 9398503. doi:10.1006/jtbi.1997.0496.
[29] Mech, L.D.; Boitani, L. 2010. Wolves: Behavior, Ecology, and Conservation. Chicago: University of Chicago Press. ISBN 0226516989.
[30] Abrantes, R. 2015. Ethology—The Study of Animal Behavior in the Natural Environment. Cambridge: Wakan Tanka Publishers.
[31] Williams, G.C. 1966. Adaptation and Natural Selection: a Critique of Some Current Evolutionary Thought. Princeton: Princeton University Press.
[32] Dawkins, R. 1976. The Selfish Gene. Oxford: Oxford University Press (1989). pp. 183–185. ISBN 0192860925.
[33] Nowak, M. A.; Tarnita, C. E.; Wilson, E. O. 2010. The evolution of eusociality. Nature. 466 (7310): 1057–1062. doi:10.1038/nature09205. ISSN 1476-4687.
[34] Wilson, E.O. 2012. The Social Conquest of Earth. Liveright. ISBN-10: 978-0871403636/0871403633.
[35] Markvoort, A.J.; Sinai, S., Nowak, M.A.. 2014. Computer simulations of cellular group selection reveal mechanism for sustaining cooperation. Journal of Theoretical Biology 357 (2014) 123–133.
[36] Wilson, E.O. 2005. Kin selection as the key to altruism: its rise and fall. Soc Res 72:159–166.
[37] Yakubu, Y. 2013. The Altruism Paradox: A Consequence of Mistaken Genetic Modeling. Biol Theory (2013) 8:103–113 DOI 10.1007/s13752-013-0120-4.
[38] Pigliucci, M. 2001. Phenotypic Plasticity: Beyond Nature and Nurture. Johns Hopkins University Press, Baltimore.
[39] Kelly, S.A., Panhuis, T.M., Stoehr, A.M. 2012. Phenotypic Plasticity: Molecular Mechanisms and Adaptive Significance. Comprehensive Physiology. pp. 1417–39. doi:10.1002/cphy.c110008. ISBN 978-0-470-65071-4. PMID 23798305.
[40] Winston, M.L. 1987. The Biology of the Honeybee. Harvard University Press, Cambridge, MA.
[41] Prete, F.R. 1990. The conundrum of the honey bees: one impediment to the publication of Darwin’s theory. J Hist Biol 23:271–290.
[42] Albert, S., Bhattacharya, D., Klaudiny, J., Schmitzova, J. & Simuth, J. 1999. The family of major royal jelly proteins and its evolution. J. Mol. Evol. 49, 290– 297.
[43] HGSC The Honeybee Genome Sequencing Consortium. 2006. Insights into social insects from the genome of the honeybee Apis mellifera.
Nature, 443, pages 931–949 (2006)
[44] Evans, J.D., Wheeler, D.E. 1999. Differential gene expression between developing queens and workers in the honey bee, Apis mellifera.
Proc Natl Acad Sci USA 96:5575–5580.
[45] Wilkinson, G.S. 1984. Reciprocal food sharing in the vampire bat. Nature 308:181–184.
[46] Packer, C. 1977. Reciprocal altruism in olive baboons. Nature 265:441–443.
[47] Wenseleers T., Ratnieks, F.L.W. 2006. Enforced altruism in insect societies. Nature 444:50.
[48] Emlen, S.T., Wrege, P.H. 1992. Parent–offspring conflict and the recruitment of helpers among bee-eaters. Nature 356:331–333.
[49] Young, J.A., Clutton-Brock, T. 2006 Infanticide by subordinates influences reproductive sharing in cooperatively breeding meerkats. Biol Lett 2:385–387.
[50] Gadagkar, R. 1997. Survival strategies: cooperation and conflict in animal societies. Harvard University Press, Cambridge, MA.
Featured illustration: Anton Antonsen.
Featured Course of the Week
Ethology Ethology studies animal behavior in its natural environment. It is one of the fundamental courses in your curriculum. A reliable knowledge of animal behavior is the basis to create a satisfying relationship with any animal we train.
Featured Price: € 168.00 € 98.00
Learn more in our course Ethology. Ethology studies the behavior of animals in their natural environment. It is fundamental knowledge for the dedicated student of animal behavior as well as for any competent animal trainer. Roger Abrantes wrote the textbook included in the online course as a beautiful flip page book. Learn ethology from a leading ethologist.